
“Was the first craniate on the road
to cognition?”
Evolution and Cognition 2003; 9(2):142-156.
Evolution and Cognition 2003; 9(2):142-156.
Fredric J. Heeren (Page 2)
|
|
||
|
|
|
|
|
|
|
|
The first generally recognized evidence
for possible chordates in the Cambrian came to light almost a
quarter of a century ago, when Cambridge paleontologist Simon
CONWAY MORRIS tentatively promoted a middle Cambrian species
called Pikaia from annelid worm to chordate status (CONWAY MORRIS /
WHITTINGTON 1979). Stephen Jay GOULD saved the tiny, simplified
eel-shaped animal for the climax of his popular book about
Canada’s Burgess Shale animals. On the last pages of Wonderful Life, he
called Pikaia “the missing and final link in our story of
contingency —the direct connection between Burgess
decimation and eventual human evolution” (GOULD 1989,
p322). Since then, however, less enthused scientists have
questioned Pikaia’s chordate classification because of its lack of
chordate features like gills, gonads, and a full notochord
(HOLLAND personal communication).
For GOULD, the middle-Cambrian Pikaia best fit what
the earliest chordate should look like: simple, sleek and
headless. He mentions no brain, eyes, or other sensory organs
when describing Pikaia in his popular book; even the possibility of a
head seems remote in an animal whose anterior end, in his
illustration, splits into two (GOULD 1989).
The next best hope came from animals whose
chordate status was disputed or who appeared too late to show
that chordates joined in the early Cambrian radiation of new
forms. The eel-like conodont, long known only from its teeth,
extended back only to the late Cambrian (PURNELL/DONOGHUE
1997). During the 1990s battles ensued over descriptions of two
new chordate claims represented by just a few specimens: Yunnanozoon (CHEN et
al. 1995)
and Cathaymyrus (SHU/CONWAY MORRIS/ ZHANG 1996). The discoverer of Cathaymyrus thought Yunnanozoon looked
more like a hemichordate (acorn worm) than a chordate
(SHU/ZHANG/ CHENG 1996); and the discoverer of Yunnanozoon opined
that his challenger had mistaken Cathaymyrus’s
squashed dorsal fin for a notochord
(CHEN/ HUANG/LI, 1999b).
After the Cambrian waters had been
sufficiently muddied, researchers wondered if any true chordate
had ever been found in Cambrian strata. Maybe our own
“sophisticated” phylum had not yet evolved. Even
GOULD’s Pikaia, though used to illustrate Cambrian chordates
in vertebrate textbooks, no longer looked convincing, since it
lacked many of the chordate features claimed by the more recent
finds.
Thus the significance of the discovery of Haikouella —displayed
in over 300 specimens. In CHEN’s description of Haikouella fossils,
he pointed out features that not only demonstrate its chordate
status, but that shed light on the origin of craniates
(biology’s new name for vertebrates) (CHEN/HUANG/LI
1999b). The new nomenclature reflects a new primary diagnostic
feature for this taxon: a distinct head enclosing a brain and
sensory organs, recognizing that this character should now take
precedence over the presence of a vertebral column.
Known for his research on amphioxus, the
present-day animal thought to best represent the ancestor of
all vertebrates, Nicholas HOLLAND said: “There’s no
question these things are chordates” (ENSERINK 1999). He
remarked on the great number of specimens with conspicuous gill
slits (for straining food out of the water) and other
diagnostic characters: “The muscle segments are
unarguable, and the notochord’s good too” (HOLLAND
1999). Unlike specimens from other recent finds, both Haikouella and Yunnanozoon exhibit
large notochords that clearly run the full length of their
bodies. “It’s the earliest known chordate
ancestor”, said HOLLAND. “Every zoology student and
every paleontology student for many, many generations is going
to have to look at that picture. This is going to be page one,
two, three and four of vertebrate texts, and paleontology
texts, and invertebrate zoology texts” (HOLLAND personal
communication).
Since the discovery of Haikouella, Degan SHU et
al. (1999) reported their discovery of two new chordates, Myllokunmingia and Haikouichthys, each
based on a single specimen. Collaborator Simon CONWAY MORRIS
proposes that the animals had skulls made of cartilage
(MONASTERSKY 1999). CHEN notes that the specimens display two
important features: distinctive fins (large dorsal and possibly
paired ventral fins) and zigzag-shaped segmented muscles,
similar to the pattern in modern fish (CHEN, personal
communication). Though paleontologists of these various
discoveries continue to contend with one another over whose
specimens are ancestral to whose—and whose are true
chordates—all agree that chordates have now been found in
the early Cambrian (ENSERINK 1999; DZIK 1995).
What will happen to GOULD’s Pikaia, the animal
zoology textbooks presently tout as our earliest chordate
ancestor? HOLLAND contends that the textbook writers had no
business picking up Pikaia as a chordate ancestor from GOULD’s
popular book, since GOULD was not an authority on the animal
(HOLLAND personal communication). GOULD had simply made it fit
what he needed to relate the Burgess Shale fauna to humans.
“Why do humans exist?” asked GOULD on the last page
of Wonderful Life. “A major part of the answer, touching
those aspects of the issue that science can treat at all, must
be: because Pikaia survived the Burgess decimation” (GOULD
1989, p323). GOULD had used Pikaia to relate Pikaia to us and us to his overriding theme:
contingency. “What this conference has done”, said
HOLLAND at the symposium where Haikouella
was announced, “is to pull
the rug out from under Pikaia, for sure. Nobody will ever talk about it
again” (HOLLAND personal communication).
Shedding Light on Vertebrate Origins
Now that lower Cambrian chordates have
been con- firmed, zoologists must deal with the fact that Haikouella —and
other early Cambrian chordates—look nothing like what
they expected to see in a predecessor of Pikaia. Rather than
finding evidence that this complex animal had less
sophisticated ancestors, CHEN and SHU instead found examples of
more complex, fully formed chordates—fifteen million
years earlier. None of these newly discovered chordates have
vertebrae or endoskeletons, so strictly speaking, they
aren’t vertebrates. But displaying relatively large
brains, these animals appear to be in the line to vertebrates,
so that at the conference where Haikouella
was announced, the strange term
“pre-backbone vertebrate” was frequently bandied
about. The brain’s early appearance would seem to
demonstrate that brain and endoskeleton did not evolve
together, as had been assumed, but rather that the brain
appeared long before the development of the vertebrate spine.
“The discovery of the first craniate
shows that the evolutionary history toward vertebrates had been
on track long before the origins of the backbone”, says
Taiwanese biologist Chia-Wei LI ( 1999), co-author of the Haikouella description.
Haikouella findings run counter to the commonly held notion that
the head could not become the dominant body structure until the
body’s superstructure was also in place. It now appears
that, against externalist expectations, cephalization (when the
head became the dominant or controlling body structure)
preceded endoskeletization (the development of an internal
support structure).
CHEN also identified other important
features in Haikouella that preceded the development of a bony skeleton:
a neural cord that, like the notocord, runs the length of the
body; a heart; a pair of lateral eyes; and tiny teeth. The
teeth are located far back in its large pharyngeal cavity
rather than in the mouth, indicating that it used them for
grinding, not biting. Biologists had assumed that chordates did
not develop the ability to accumulate minerals in their bodies
to form teeth or bones until about 500 million years ago. But Haikouella and Yunnanozoon demonstrate
that biomineralization had begun at least 30 million years
earlier. Teeth led the way long before the development of a
notochord-protecting, mineralized vertebral column or other
bones.
Constraints, Channeling, and Convergence
The sudden explosion of widely disparate
Cambrian animal Bauplans, followed by no new body plans
throughout the rest of geologic history, fits the picture of a
constrained process, the channeling of changes within
particular forms. Scientists also find evidence of constraints
today in the form of parallelism and convergence,9 both in
experiments with living animals and in theoretical modeling.
From his research on the development of amphibians, brain
researcher Gerhard SCHLOSSER notes trends “where several
characters tend to act as a ‘unit of evolution’,
i.e., they tend to coevolve repeatedly” (SCHLOSSER 2000).
Evolutionary geneticist Paul RAINEY and
his colleagues have also noticed convergence in evolution while
experimenting with the bacterium Pseudomonas
fluorescens. “These
experiments in test-tube evolution”, says RAINEY,
“allow us to replay life’s tape, albeit on a small
scale, as often as we like” (RAINEY 2003). Their
findings? “Evolution repeats itself”. By growing
rapidly diversifying strains of the bacterium in test tubes of
nutrient broth, they have discovered that “in the face of
similar selective conditions, different lineages can find
similar solutions to the same problems”. RAINEY is not
afraid to find implications from his findings for human
evolution: “Replay life’s tape”, he claims,
“and while Homo sapiens may not evolve there is a high probability that
introspective bipedal organisms with binocular vision
will” (Ibid).
Simon CONWAY MORRIS reaches a similar
conclusion. Speaking of the property of consciousness, he
writes: “Here the reality of convergence suggests that
the tape of life, to use GOULD’s metaphor, can be run as
many times as we like and in principle intelligence will surely
emerge” (CONWAY MORRIS 1998, p14). What about “the
numerous entirely plausible alternatives of strikingly
different forms” that GOULD expected if the tape should
be rerun from the beginning? “Put simply”, says
CONWAY MORRIS, “contingency is inevitable, but
unremarkable…. There are not an unlimited number of ways
of doing something. For all its exuberance, the forms of life
are restricted and channeled” (p13). CONWAY MORRIS
believes that convergence “effectively undermines the
main plank of GOULD’s argument on the role of contingent
processes in shaping the tree of life” (Ibid). GOULD, he
says, “presupposes that constraints are weak” and
makes a “most egregious misinterpretation of the Burgess
Shale” (CONWAY MORRIS 1998–1999). His
“egregious misinterpretation” —contingency as
the major lesson of the Burgess Shale—is a conclusion
that GOULD drew from his personal credo, according to CONWAY
MORRIS, not from paleontology (Ibid).
Hierarchies
Cladistics, a branch of biology that does
indisputably draw its evidence from paleontology, hypothesizes
relationships between organisms according to shared derived
characters (synapomorphies). The distribution of these
diagnostic features forms a set of nested groups (clades), in
which smaller clades are contained within larger ones. The
hierarchic pattern that has become the hallmark of cladistic
analysis is related to the lack of transitional forms found
between groups. DARWIN expected evolution to leave us with
surviving modern groups within groups, but he expected the history of life to
proceed in a gradualistic sequence that blurs the lines between
groups. The scarcity of such fossil transitions can only be
explained in DARWINIAN terms as a sampling problem, an artifact
of an incomplete fossil record (DARWIN 2000, p292). Modern
paleontologists generally agree, however, that the fossil
record is actually robust enough to tell us that the scarcity
of transitional forms is real and significant (SIMPSON 1960;
GOULD 1977; VALENTINE/ERWIN 1987; DONOVAN/ PAUL 1998; FOOTE
1996; FOOTE/SEPKOSKI 1999), making the hierarchic pattern a
genuine aberration in the gradualistic picture.
The priority of typology over continuity
has persisted, according to SIMPSON, among “all schools
of taxonomy including some that usually oppose typology in
principle” (SIMPSON 1961, p49). Haikouella contributes to
this crystalizing picture of distinct, fully formed body plans
from near the start. Developmentalists observe the same
hierarchical processes at work in both ontogenesis and
evolution. Biologist Brian GOODWIN writes: “Developmental
processes are hierarchical. So are biological classification
schemes” (GOODWIN 1994, p234). Wallace ARTHUR agrees:
“A theme running through the work of most contributors to
what can now be described as evolutionary developmental biology
is the relationship between these two hierarchies”,
(ARTHUR 1997, p256) and he asserts that “it is informative about
the nature of evolutionary mechanisms” (p257).
Saltation
How much further back can we trace our
ancestors? Nicholas HOLLAND, for one, wants to know what preceded these
complex, early Cambrian craniates, a question, he says, that
remains as big a mystery as ever: “Where are Haikouella’s ancestors?
The sixtyfour dollar question is, What is this hooked to? That
nobody knows” (HOLLAND personal communication).
In his presentation to an international
symposium on Cambrian body plans (1999), HOLLAND gave genetic
reasons why the most popular theoretical predecessor for
chordates, tunicates (sea squirts), only works in the
imagination of the theorists. When chordates are compared
genetically with tunicates and fruit flies, he says, “the
fruit fly is closer to the tunicate every time” (HOLLAND
personal communication).
No obviously ancestral fossils presently
exist to support theories about how chordates, or the other
phyla, evolved in Precambrian times. “There are a lot of
different totally cutup paper doll ideas about where things
come from that aren’t based on fossils at all, but people
sitting in their armchairs”, says HOLLAND (personal
communication). The ceaseless re-interpretation of ancestral
lineages for the phyla is easily demonstrated by the relevant
literature (ARTHUR 1997, p73; BERGSTRÖM 1994; LYNCH 1999).
Wherever the first chordates came from, HOLLAND thinks science
must now take seriously the concept of “saltation”,
the possibility of evolution in quick jumps. However broadly
one defines “saltation”, paleontological evidence
for the notion is certainly supportive of the
internalist/developmentalist position.
Though opinions vary about the Precambrian
antiquity of the phyla, all agree that almost all of these most
widely separated animal groups had appeared by the early
Cambrian period. Why didn’t new phyla continue to evolve
during subsequent eras? Why did such disparate phyla as
chordates, mollusks, arthropods, and the 35-or-so others first
show up in the fossil record so close to the same time? CHEN
places the window of opportunity for the explosive evolution of
the majority of body plans within a narrow window of three
million years (CHEN 1999), though of course, this is hotly
disputed.
Body plans seen in the Precambrian include
sponges, annelid worms, and echinoderms (like sea stars), but
little else to represent the many lineages expected to lead to
the 35 Cambrian groups. Gradualists have claimed that the
ancestors of the many other disparate Bauplans must have been
too small or too soft to be preserved. But since 1998,
phosphate deposits at a Precambrian locale called Weng’an
have proved capable of preserving the smallest and softest
organisms imaginable (LI et al. 1998). Sponge embryos have been
found by the thousands in early cleaving stages, seen under the
microscope in groups of 2, 4, 8 cells, etc. (Figure 3). Though
small and soft specimens are found in abundance, the number of
body plans remains small.
The questions raised by such findings drew
sixty scientists to Kunming, China, for a symposium entitled:
“The Origins of Animal Body Plans and Their Fossil
Records”. Perhaps it took the discovery of our own
phylum’s participation in the early Cambrian big bang to
bring together such an international gathering to consider a
pattern some call “top-down evolution”.
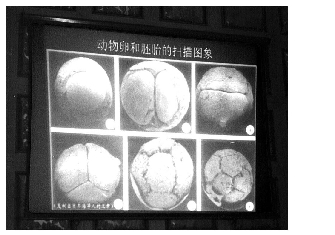
Figure 3. Sponge
embryos seen under the microscope at the cellular level in
early cleaving stages, well preserved by the thousands from
Precambrian deposit at Weng’an.